今天介绍一篇由Sarah E. Berry等人前段时间发布于nature medicine的一篇文章。文中对英国(n=1002)和美国(n=100)的年轻健康成年人进行餐后代谢反应评估,并开发机器学习模型来预测人体内的甘油三酸酯(r=0.47)和血糖(r=0.77)对食物摄入的反应,这项技术有助于制定个性化的饮食策略。
1. 介绍
现有的营养研究和相应的指南均着重于人群平均值,但人体对食物的反应却有高度的特异性,这很可能是因为来自于每个人独有的生物学特征(如菌群和核DNA的差异)及生活方式。尽管多数临床诊断中使用空腹血液测定法测血糖,但大多数人在白天处于餐后状态的。餐后血脂、葡萄糖和胰岛素稳态失衡是营养相关的非传染性疾病(NCDs)和肥胖的独立风险因素。
对饮食组成的个体反应(PREDICT 1)临床试验(NCT03479866)旨在定量和预测个体对标准膳食的餐后甘油三酯、葡萄糖和胰岛素的差异。作者招募了来自英国的1002名成年人(包括非双胞胎、同卵双胞胎和异卵双胞胎)并获得遗传、代谢、菌群、膳食成分和膳食背景数据以区分出个体膳食反应的预测因子,然后预测在美国无亲缘关系的成年人中得到验证。
作者研究发现人们之间,同卵双胞胎之间的餐后反应差异很大,这在很大程度上归因于可变因素(modifiable factors)。此外,对某一特定食物代谢反应较差的人,对具有相同常量营养素的其他食物的代谢反应也可能较差。研究还得出餐后葡萄糖和甘油三酯反应之间的总体相关性很弱。此次试验也验证了作者开发的餐后预测模型可以帮助优化个性化饮食建议。
2. 结果
作者从英国组的1002名健康成年人收集基线临床测量值,包括连续混合营养饮食的餐后代谢反应(0-6h,血液中甘油三酸酯、葡萄糖和胰岛素的浓度)。受试者随后进行13天居家实验并收集其他数据:使用连续血糖监测仪(CGM)和干血斑(DBS)分析测量了8种(meal 7重复)不同常量营养素(脂肪,碳水化合物,蛋白质和纤维)含量膳食的餐后反应。研究设计见Fig.1。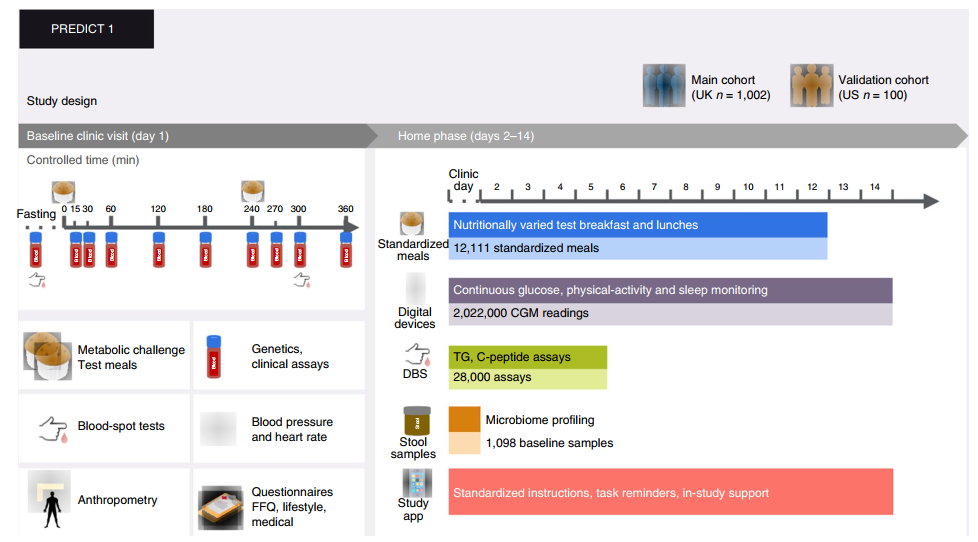

Fig.1 PREDICT 1研究包括英国受试组(nmax=1002名受试者)和一个美国验证组(nmax=100名受试者)。TG,甘油三酸酯。
2.1 餐后反应的个体间和个体内差异
作者在严格控制的临床环境中进行空腹后的连续标准膳食测试,研究了餐后反应的个体反应(见Fig.2a)。使用Levene’s方差检验评估每个结果的个体间反应模式。实验结果表明:餐后甘油三酯和葡萄糖浓度比空腹值能更好的区分个体的代谢耐受性。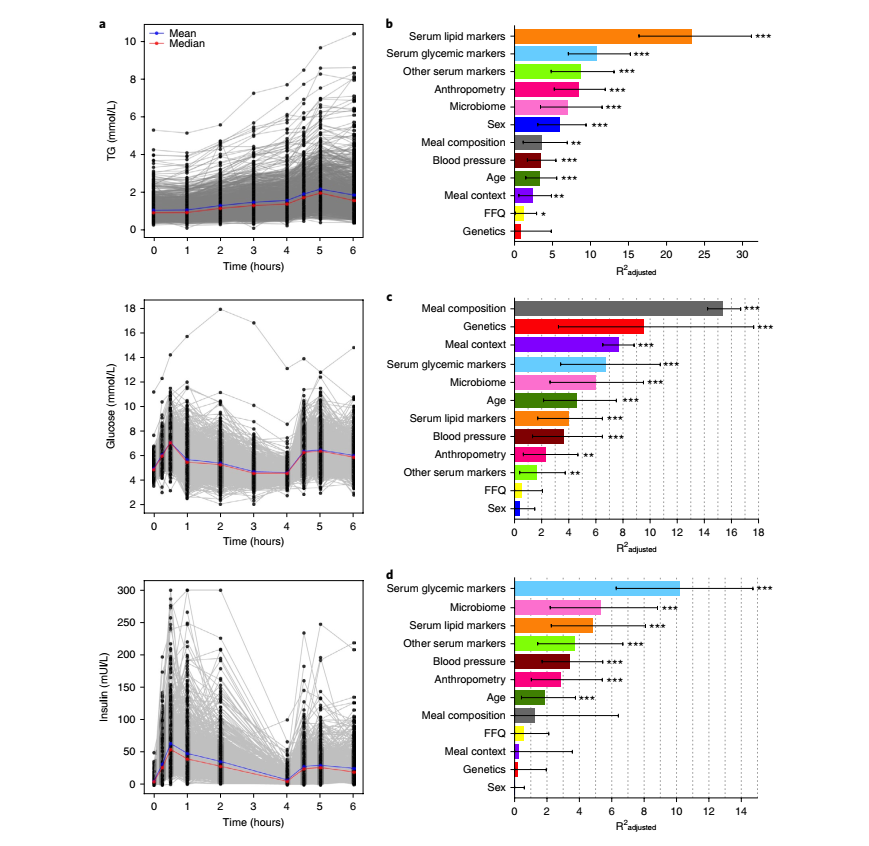
Fig.2 餐后反应的差异
a. 在临床中,早餐和午餐的餐后反应,甘油三酯、葡萄糖和胰岛素的个体间差异(n=1002)。b. DBS测定甘油三酯6h-rise的决定因素(比较meal 1和meal 7)。c. 用CGM测定葡萄糖iAUC0-2h的决定因素(比较7种膳食;1,2,4,5,6,7,8)。d. 使用DBS分析作为替代胰岛素的C-peptide1h-rise的决定因素(比较meal 2和meal 3)。每个输入变量的特征方差解释来自独立的(非遗传)回归模型。数值代表调整后的方差解释比例(R2),误差棒的CIs为95%。根据膳食成分和膳食背景调整的R2值来自以下膳食样本量:甘油三酯6h-rise,n=712;葡萄糖iAUC0-2h,n=9102;C-peptide1h-rise,n=186。所有其他决定值来自以下膳食样本量:甘油三酯6h-rise,n=920;葡萄糖iAUC0-2h,n=958;C-peptide1h-rise,n=960。*P < 0.05,**P < 0.01,***P < 0.001,使用多元线性回归。
作者在开发个性化预测算法时,一个关键的假设是 :个体对相同膳食的独特反应是可重复的。在单一测试反应情景对饮食干预反应的研究中观察到的大多数个体内的表型差异性,可能是趋均数回归或是其他误差导致的。因此在相似的条件下,给每个参与者两份不同常量营养素成分的测试餐。
2.2 预测人群中个体的餐后反应
作者评估了输入变量预测个人餐后反应的总体程度(Fig.2b-d),首先使用多变量线性回归。输入变量包括:(1)基线特征(年龄、性别、临床生化(血脂、血糖和其他测量)、人体测量学);(2)遗传学(单核苷酸多态性(SNPs));(3)消化系菌群特征;(4)习惯性饮食(从食物频率问卷(FFQ));(5)用餐背景(睡眠、前一餐、体育活动、用餐顺序和/或时间);(6)膳食组成(能量来自碳水化合物、糖、脂肪、蛋白质和纤维)。餐后血糖反应由临床和居家阶段的血清和CGM测量值来确定(7种标准膳食)。餐后C-peptide和甘油三酯的测定(2种标准膳食)由血清和DBS测定在临床和居家阶段收集的材料确定。还测试了空腹特征和餐后特征之间的相关性,发现餐后甘油三酯、血糖和C-peptide测量值之间的相关性较低(Fig. 3a)。
个体基线特征:个体基线特征解释的甘油三酯6h-rise、葡萄糖iAUC0-2h和C-peptide1h-rise特征方差比例见Fig. 2b-d。
遗传因素:对230对双胞胎(n=460;183个同卵和47个异卵)的遗传性进行研究。可积累遗传因素解释了葡萄糖iAUC0-2h中48%的方差,甘油三酯6h-rise0%的方差和胰岛素2h-rise9%的方差(Fig. 3b)。胰岛素1h-rise和C-peptide1h-rise的遗传方差接近0。
SNP-based遗传因素:一部分双胞胎受试者进行了全基因组基因分型,已有全基因组关联研究(GWAS)数据(n=241)。一些SNPs与这些变量显著相关(P<0.05)(Fig. 3c),但它们共同解释的葡萄糖iAUC0-2h观察到的差异仅占9%(Fig. 2c),而餐后甘油三酯和餐后C-peptide的方差不足1%(Fig. 2b,d)。
肠道微生物组(16S核糖体RNA):不对任何其他个人特征进行调整时,微生物组解释餐后甘油三酯6h-rise的7.5%,餐后葡萄糖iAUC0-2h的6.4%的和餐后C-peptide1h-rise的5.8%。
膳食构成、习惯饮食和膳食背景:确定食物的常量营养素组成的影响,对一部分受试者测量甘油三酯6h-rise和C-peptide1h-rise两种不同常量营养素成分的居家阶段标准膳食(甘油三酸酯,meal 1和7;对于C-peptide,meal 2和3)(n=712和n=186)。葡萄糖iAUC0-2h在7种标准膳食中测量(meal 1、2、4、5、6、7和8),总计920个人,9102餐。Fig. 2b、c、d显示了由膳食组成、习惯饮食和膳食背景解释的甘油三酯6h-rise、葡萄糖iAUC0-2h和C-peptide1h-rise的差异比例。多变量回归模型(meal 1、2、4、5、6、7和8)显示,调整碳水化合物消耗后,每1g脂肪、纤维和蛋白质,葡萄糖iAUC0-2h(mmol per l per s)显著(P<0.001)减少79、142和185。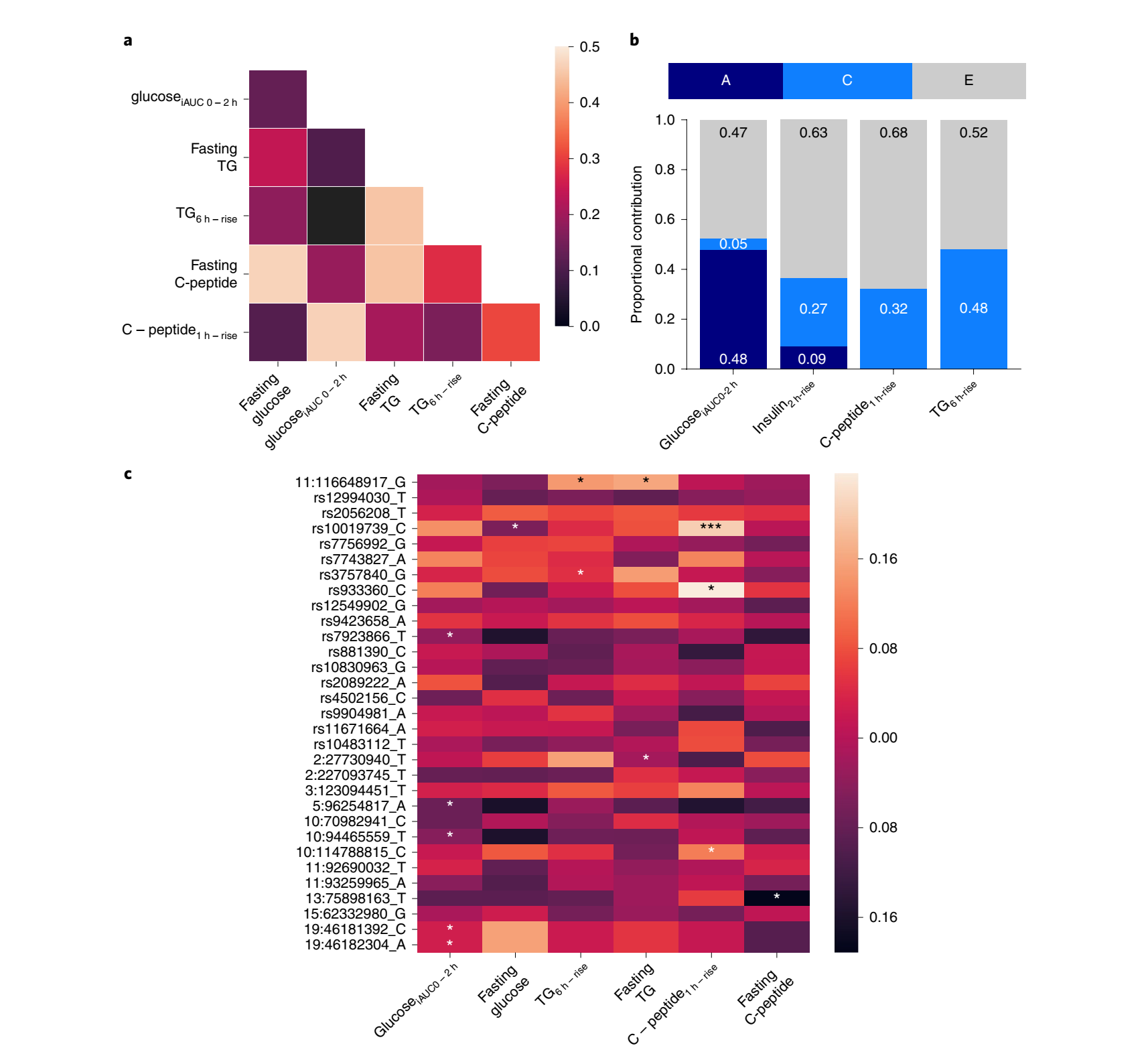
Fig.3 基线值、遗传和微生物组因素与餐后反应的关系。a.来自英国组的980名受试者的基线值和餐后预测测量之间的Pearson相关性。b.在183对同卵双胞胎和47对异卵双胞胎中,餐后反应的遗传性(ACE模型对甘油三酯、葡萄糖、胰岛素和C-peptide的对数标尺餐后反应进行拟合)。在ACE模型中,A表示可加性遗传成分;C共享的环境成分;以及个体的环境因素。c.SNP与餐后测量的相关性,主要集中选择的餐后特征GWASs中确定的SNP(n=241;*P < 0.05,***P< 0.001,使用双面卡方检验)。
机器学习模型:作者为了估计所分析因素的无偏预测效用,使用了对过拟合具有稳健性的机器学习方法。随机森林回归模型使用英国组数据集的所有信息特征(膳食组成、习惯饮食、膳食背景、人体测量、遗传学、微生物组、临床和生化参数)来预测甘油三酯6h-rise、葡萄糖iAUC0-2h和C-peptide1h-rise。对每个特征利用皮尔逊相关系数(r)将预测值与测量值进行比较;甘油三酯6h-rise、葡萄糖iAUC0-2h和C-peptide1h-rise的相关性分别为r=0.47、r=0.77、r=0.30。在held-out法验证集中(美国组)中观察到类似的相关性。模型预测甘油三酯6h-rise和葡萄糖iAUC0-2h分别为r=0.42和r=0.75,而C-peptide1h-rise的预测较差(r=0.14)(Fig. 4)。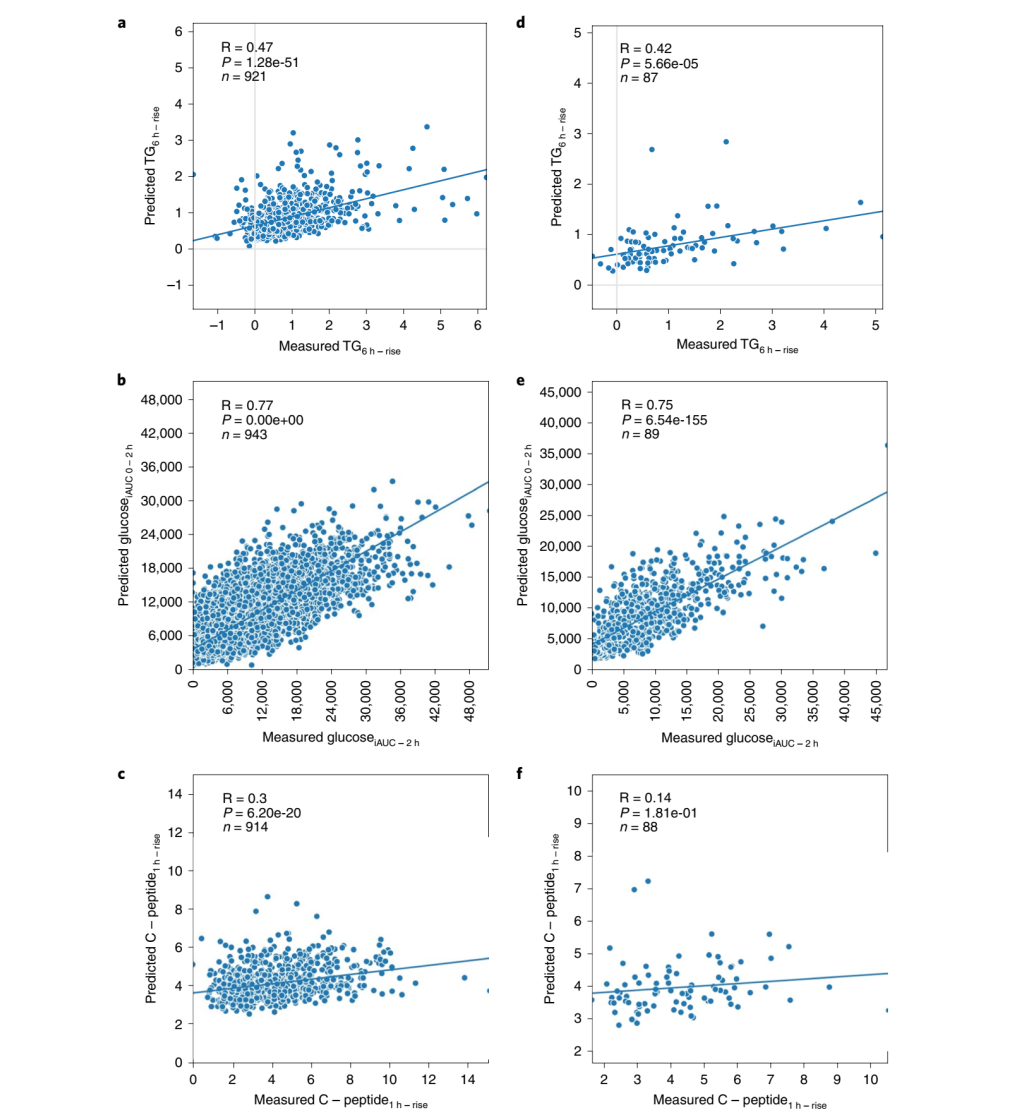
Fig.4 机器学习模型拟合餐后测量值。a-c,来自英国组中甘油三酯6h-rise(a),葡萄糖iAUC0-2h(b)和C-peptide1h-rise(c)餐后反应的机器学习模型的结果。d-f,机器学习模型应用于美国验证组的结果。变量之间的关系用Pearson’s相关系数(r)表示,绘制回归线;n表示受试者人数。预测每个值所使用的特征与Fig. 2b-d中的线性模型所列出的特征相同。
餐后反应与临床结果替代评分的关系:作者比较了不同生物标记物在空腹和餐后的含量,用于通过比较受试者工作特征曲线(ROC-AUC)下面积(Fig. 5),预测糖耐量受损(口服糖耐量试验(OGTT)后2h 7.8-11.0mmol l-1)和动脉粥样硬化性心血管疾病(ASCVD)10年风险评分。结果得出空腹甘油三酯和甘油三酯6h-rise对ASCVD风险的ROC-AUC的占比相似,且把两者都包括在内,比只包括其中一个更有价值(Fig. 5a)。作者还发现,尽管餐后血糖的信息不如空腹血糖有价值,但在空腹血糖中添加葡萄糖iAUC0-2h可使ASCVD10年风险评分的ROC-AUC略微提高(0.72vs0.69)。空腹C-peptide和空腹血糖与空腹甘油三酯在预测ASCVD方面同样有效(ROC-AUC=0.69),而餐后C-peptide(ROC-AUC=0.63)和餐后血糖(ROC-AUC=0.62)弱于餐后甘油三酯(ROC-AUC=0.71)。空腹和餐后甘油三酯浓度对糖耐量受损(IGT)的预测能力较弱(ROC-AUC分别为0.55和0.59),而空腹和餐后C-peptide浓度对糖耐量受损(IGT)的预测能力中等(ROC-AUC分别为0.64和0.65),合并使用没有增加预测价值。作者未利用CGM的葡萄糖数据预测IGT。这是因为IGT仅仅是根据OGTT期间2小时的血糖浓度来定义的,该血糖浓度由CGM葡萄糖记录收集,因此预测因子和临床评分变量的推导在很大程度上依赖于彼此。英国和美国组的结果相似(Fig. 5)。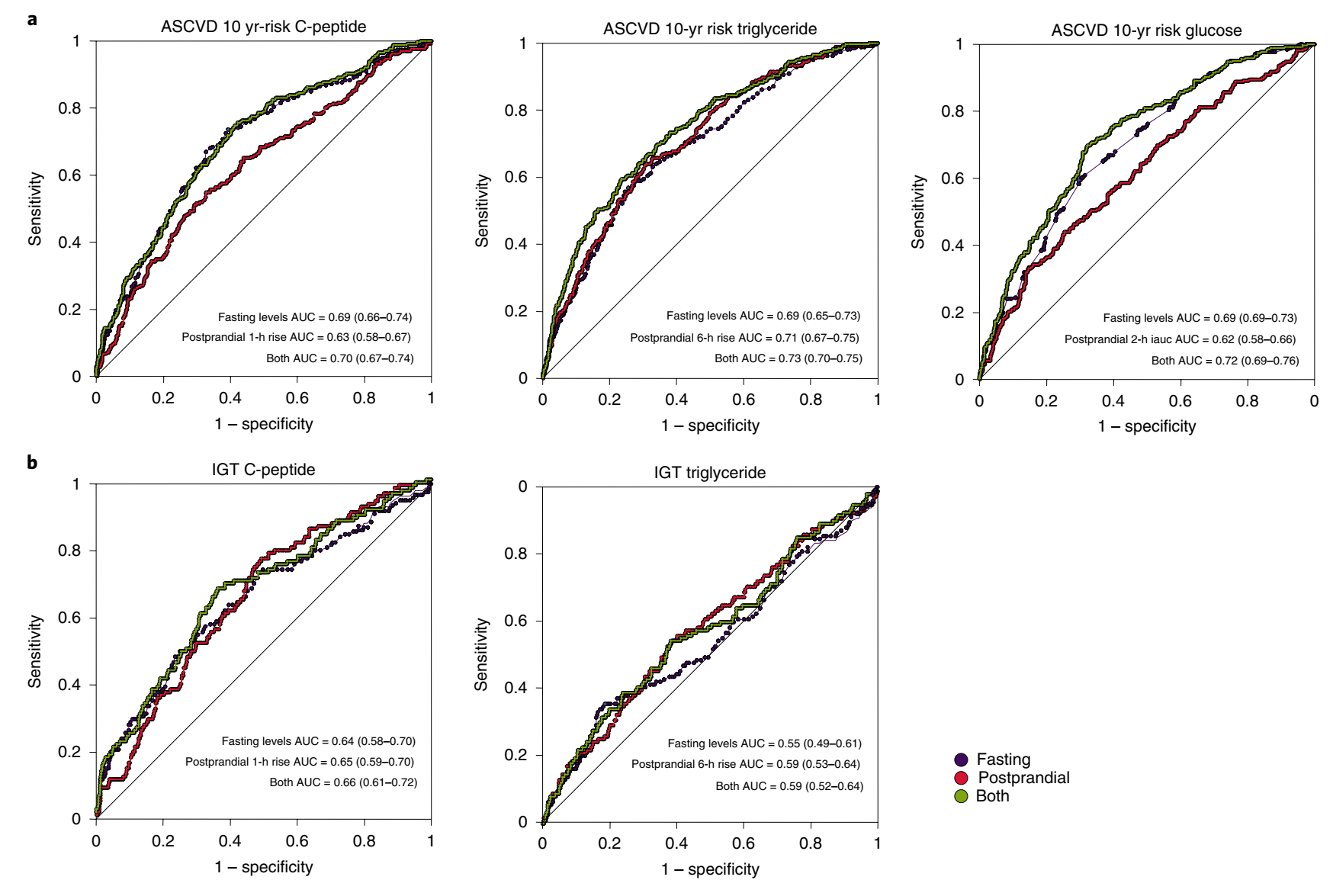
Fig. 5 英国组临床测量中空腹和餐后甘油三酯、C-peptide和葡萄糖浓度的关系。a、b显示禁食和餐后甘油三酯、葡萄糖和C-peptide测量值预测效用的ROC曲线,以区分底部的70%和顶部的30%(ASCVD10年风险界值,0.0183)ASCVD10年风险(n=951,英国组)(a)和糖耐量受损(IGT)(n=826,英国组)(b)同样的分析在美国组(n=92)导致ROC-AUC ASCVD10年期(95%置信区间)值的风险:C-peptide空腹AUC=0.68(0.56-0.80),餐后AUC=0.66(0.54-0.77),AUC=0.69(0.58-0.81);甘油三酯 空腹AUC=0.73(0.63-0.84),餐后AUC=0.75(0.65-0.85),both AUC=0.77(0.67-0.88);葡萄糖 空腹AUC=0.74(0.63-0.85),餐后AUC=0.64(0.52-0.76),both AUC=0.76(0.64-0.85)。对于IGT,值为:C-peptide 空腹AUC=0.66(0.53-0.80),餐后AUC=0.59(0.46-0.72),both AUC=0.67(0.54-0.80);甘油三酯 空腹AUC=0.66(0.53-0.80),餐后AUC=0.59(0.46-0.72),both AUC=0.61(0.54-0.80)。
2.3 解读个体反应
作者表示在居家阶段,由于甘油三酯和C-peptide无法收集到精细数据,故基于CGM数据,研究个体的餐后反应在多大程度上归因于一顿饭的血糖特性,并将其与其他可变因素(如用餐时间、运动和睡眠)的变化进行了比较。
作者探索了是否存在对个人而言独特的膳食特定反应。将每个参与者的葡萄糖iAUC0-2h对每一对可能的标准膳食排序。然后确定了这些排名对每个参与者的相异频率。对于大多数餐组,所有个体的排名是相同的(Fig. 6a)。然而,对于所选的膳食对,高达48%的受试者的排序相异。使用方差分析(ANOVA)来估计解释血糖反应的不同因素的效应大小(Fig. 6b),包括特定人的影响(不同人之间有差异,但不同膳食之间没有差异)。文中不仅考虑膳食常量营养素和能量含量对反应的影响(膳食组成),还考虑相对于人群,每个人对所有固定餐的平均反应如何(个人葡萄糖量表),以及个体特定饮食反应的影响,误差可归因于葡萄糖测量和其它来源的变化(包括可改变的变化来源,如睡眠、昼夜节律和运动)。
作者发现,与前面描述的线性模型一致,方差分析模型显示,有三个与膳食相关的因素解释个体的血糖反应。膳食常量营养成分改变iAUC 16.73%(CI 15.37%-18.92%),但个体葡萄糖比例更大,iAUC改变约18.74%(17.96%-19.46%)。个体的膳食特异性反应要小得多,影响最后一餐的iAUC 7.63%(6.11%-8.96%)。其他不直接与膳食构成相关的可改变的变化来源,如用餐时间、运动和睡眠,贡献了与膳食构成类似的方差(Fig.6 b、c)。
作者为了调查改变三餐的顺序和时间是否会影响血糖反应,观察了早餐和午餐相同(meal 2)的受试者。同个体午餐(葡萄糖iAUC0-2h=14254,s.d.=6593)(早餐后4h)摄入的血糖反应平均比早餐(葡萄糖iAUC0-2h=7216,s.d.=4157)摄入的血糖反应平均高两倍(t=-35.7,2721 d.f.;P<0.001),尽管个体间差异很大(Fig. 6c)。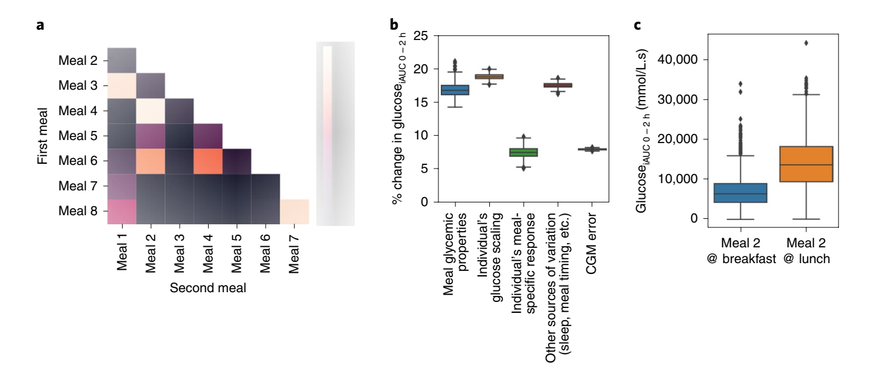
Fig.6 餐后反应的个体特异性多样性。a. 在PREDICT 1研究中,对固定膳食的血糖反应(葡萄糖iAUC0-2h)排序被改变的次数比例(n=828,英国组)。b. 解释血糖反应的因素的效应大小。使用双因素方差分析估计了不同的变异来源(n=483)。c. 同一餐后一天中时间对血糖的影响。(n=920,英国组)。箱形表示四分位数(25、50、75百分位数),95%置信区间。
3. 讨论与结论
本文作者进行了一项为期2周的干预性试验,包括临床1天和13天的居家阶段。在该试验中,获得了来自英国和美国的1000多名健康成年人对一系列标准膳食的餐后代谢反应。主要目的是推导出一种算法,来预测一个人对特定食物的餐后代谢反应。核心结果是甘油三酯、葡萄糖和胰岛素/C-peptide在血液中浓度的变化,因为这些生物标记物协同作用影响心脏代谢风险。
与预期相反,基因并不是大多数这些反应的主要决定因素。作者发现,餐后甘油三酯(6h-rise,0.0%)、C-peptide和/或胰岛素浓度在1小时(0.3%)和2小时(9.1%)时的遗传分数(由可加性遗传因素解释的特征方差)较低,餐后血糖(2h-iAUC)反应的遗传分数相当高(48%)。而可改变的因素,如用餐时间,比预期的影响更大。
作者发现膳食组成和背景(例如,进餐时间、运动、睡眠和昼夜节律)是餐后代谢的核心决定因素。这些预测通过肠道微生物组多样性的数据得到了加强。利用结合所有相关数据的机器学习,可以有意义地预测个体的餐后甘油三酯和血糖反应。餐后甘油三酯和葡萄糖浓度可用于测定IGT和心血管疾病风险。然而,餐后C-peptide浓度没有提供比空腹浓度更多的信息。
尽管已经开发出了这些预测算法,但仍有改进的空间,如纳入更多样化的膳食干预,以及比当前研究中使用的更详细的环境因素评估。技术进步也有助于改善预测。例如,虽然可以用CGMs连续评估葡萄糖,但目前还没有适合评估胰岛素和甘油三酯浓度的家用设备。此外,由于膳食碳水化合物与脂肪在耐受性和可检出限上的差异,餐后葡萄糖的预测在方法上优于甘油三酯反应(Fig.2b-d)。局限性在于难以直接比较甘油三酯和葡萄糖的变化。由于减少了模型误差和研究非线性响应模式的能力,对这些特征进行连续、准确的测量可以极大地改善预测。深度生物组学数据的加入可以进一步增强这些算法的预测能力;例如,使用来自16S rRNA测序的微生物组数据对预测很有价值(分别解释葡萄糖和甘油三酯反应的6.4%和7.5%的方差),但如果来自更高分辨率的宏基因组测序,数据可能会提供更多信息。在算法中加入更大的代谢组群,例如元转录组、转录组或蛋白质组,会增加成本,但也会增强预测。
研究发现习惯饮食解释了小部分(<2%)个体的餐后反应。然而FFQs有众所周知的局限性。与短期评估相结合,比如PREDICT study应用程序中包含的称重饮食记录,可能有助于缓解这些限制。本文中使用了6小时餐后测试,因为这被认为是大多数受试者可能接受的最长时间。较长时间(高达8小时)的试验数据可提供葡萄糖和甘油三酯反应方面的更有价值的信息。对于餐后甘油三酯和葡萄糖反应,英国组得出的预测模型在美国验证组中表现得几乎一样好,考虑到环境因素的差异,模型是可靠的;然而这两组都是由欧洲血统的年轻健康成年人组成的。因此,模型需要在非欧洲血统的人、老年人和有影响新陈代谢疾病的人(如糖尿病)中进行验证。
这些发现,除了评估方法的可扩展性和这里描述的预测算法的准确性,意味着至少从心脏代谢健康的角度来看,全民个性化营养具有作为疾病预防策略的潜力。
参考文献
- Sarah E. Berry,Ana M. Valdes,David A. et al. Human postprandial responses to food and potential for precision nutrition[J]. Nature Medicine,2020,26(6).
关注我们
原创文章,作者:ifyoung,如若转载,请注明出处:https://www.drugfoodai.com/ai-nutrition.html
注意:本站原创指的是原创编译,并不主张对所介绍的工作的版权,如有侵权,请联系删除!
 微信扫一扫
微信扫一扫  支付宝扫一扫
支付宝扫一扫 相关推荐
-
湖南大学吴海龙课题组:高效液相色谱指纹图谱结合化学计量学的白术有效成分分析及产地鉴别研究
该研究提出运用高效液相色谱-二极管阵列检测器(HPLC-DAD)结合化学计量学方法快速分析不同地区白术中12种活性成分含量并进行产地判别。首先,使用HPLC-DAD结合二阶校正算法(ATLD)对来自不同地区的白术中12种活性成分同时进行定性定量分析。利用“二阶优势”,12种化合物不仅在12.5 min内快速洗脱,而且还在色谱峰高度重叠的情况下实现了准确定性定量(平均回收率为80.8–109.9%),一系列品质因子参数均反映出所提方法的可行性。基于ATLD解析所得的12种活性成分和31种未校正成分的相对浓度值,使用偏最小二乘-判别分析法(PLS-DA)对不同地理来源的白术样本进行判别分析,三个地区的白术都有明显的聚类趋势,测试集的正确分类率可达90%。变量重要性投影(VIP)分析结果表明,紫丁香酚苷、白术内酯Ⅲ、白术内酯Ⅰ和苍术酮可作为白术产地判别的主要标志成分。一系列结果均证明了所建立模型的可靠性。该方法的建立有助于白术的临床使用和市场监管。
-
人工神经网络在食品加工研究中的文献调查
人工神经网络(ANN)在各个应用领域的使用水平日益提高,因其能够解决许多应用领域中的复杂问题而广受欢迎。今天给大家介绍一篇由Janmenjoy Nayak等人发表在Computer Science Review上的一篇综述,该文详细分析了人工神经网络在食品加工中的应用进展。
-
INT J PROD:影响人工智能在食品供应链中应用的关键成功因素
今天介绍一篇由英国布鲁内尔大学Manoj Dora等人于2021年8月在线发表在International Journal of Production Research上的文章。该研究旨在通过文献综述,确定人工智能在食品供应链中应用的关键成功因素。
-
食品质量和真伪分析评估的数据挖掘/机器学习方法
近年来,为了更好地鉴定食品,通过现代分析仪器所获得的数据种类和数量急剧增加。一些模式识别工具已经被开发来处理大量复杂的有效试验数据。应用最广泛的方法有主成分分析(PCA)、部分最小二乘判别分析(PLS-DA)、类模型方法(SIMCA)、k-最近邻分类算法(kNN)、平行因子分析(PARAFAC)和多元曲线分辨率-交替最小二乘分析(MCR-ALS)。然而,也有一些替代的数据处理方法,如支持向量机(SVM)、分类回归树(CART)和随机森林(RF)等,与传统的数据处理方法相比,显示出巨大的潜力和优势。在这篇文章中,作者解释了这些方法的背景,并回顾和讨论了这三种方法在食品质量和真实性领域的应用研究的报道。此外,作者声明清楚了在这一特定研究领域中使用的专业术语。
-
FOOD CHEM|海南大学云永欢课题组:高光谱成像技术结合数据融合的罗非鱼鱼片新鲜度快速检测研究
该文研究了两种波段范围的高光谱成像系统(可见-近红外光谱(Vis-NIR)和近红外光谱(NIR))在冷藏期间测定罗非鱼鱼片中挥发性盐基氮(TVB-N)含量的潜力。利用Vis-NIR和NIR数据,建立了高光谱图像中罗非鱼鱼片平均光谱与其TVB-N含量之间的校正模型,并采用数据融合和多种变量选择方法对模型进行优化。最后,采用优化的模型来实现罗非鱼鱼片中TVB-N含量的可视化分布。结果表明,高光谱成像技术结合数据融合和变量选择等化学计量学方法在罗非鱼鱼片新鲜度无损评价分析中具有可行性。
-
一种用于辨别中国白酒香气的仿生嗅觉识别系统
今天介绍一篇由北京汉王科技有限公司智能感知实验室的Weihong Liu等人于2022年3月在线发表在Food Chemistry上的文章。本文受小鼠嗅觉系统的启发,开发了一种仿生嗅觉识别系统来区分复杂的气味刺激,将气味受体(OR)的非凡特异性与反向传播(BP)神经网络的出色数据处理能力相结合,以区分中国白酒的香气。
-
Nutrients:eBASIS数据库中的可提取和不可提取抗氧化剂成分:人类健康和疾病研究中膳食评估的关键工具
今天介绍一篇由Jenny Plumb等人于2020年11月在线发表在Nutrients(IF=6.71)上的文章。这篇文章讨论了eBASIS数据库的更新,该数据库包括可提取和不可提取的抗氧化剂,这对于评估食物基质的抗氧化性质及其与人类健康的相关性非常重要。文章还介绍了非可提取多酚的重要性、测量食物和膳食补充剂中抗氧化活性/容量的各种方法、维护和更新食物成分数据库的重要性以及相关挑战。总体而言,文章强调了需要标准化的方法和数据库来准确评估我们饮食中的生物活性化合物及其潜在的健康益处
-
使用可解释人工智能(XAI)技术解开送餐服务评论的深度学习模型
今天介绍一篇由悉尼科技大学土木与环境工程学院工程与信息技术学院高级建模和地理空间信息系统中心(CAMGIS)的Anirban Adak等人今年七月发表于Foods(IF: 5.561)的一篇文章。该研究通过比较食品配送服务(FDS)领域中的简单和混合深度学习(DL)技术(LSTM、Bi-LSTM、Bi-GRU-LSTM CNN)进行了情绪分析,并使用SHapley Additive exPlanations(SHAP)和Local Interpretable Model-Agnostic Explanations(LIME)解释了预测。DL模型在从ProductReview网站提取的客户评论数据集上进行了训练和测试。结果表明,LSTM、Bi-LSTM和Bi-GRU-LSTM-CNN模型的准确率分别为96.07%、95.85%和96.33%。LSTM模型相比其他两个DL模型实现了更低的假阴性率。可解释人工智能(XAI)技术,如SHAP和LIME,揭示了用于验证模型的单词对积极和消极情绪的特征贡献。
-
预测下一场革命:食品技术对消费者接受度和满意度的影响
撰文和翻译:王雪洁 编辑:肖冉 今天介绍一篇由Nuria Recuero-Virto等人于2022年1月在线发表在British Food Journal上的文章。本文是通过收集和…
-
基于多元素筛选的化学计量学和创新多维数据分析的意大利牛肝菌产地溯源
“食品原产地认证”和“食品欺诈”一直以来都是令人感兴趣的话题。今天介绍一篇由Antonio Francesco Mottese等人发表于Food Control 的文章。该研究基于来自意大利、中国和波兰采集的牛肝菌样品的元素特征,结合主成分分析(PCA)、逐步典型判别分析(stepwise-CDA)、分类回归树(CART)、偏最小二乘判别分析(PLS-DA)和创新多维分析(MDA)建立统计模型,通过标准化学计量学技术和创新化学计量学技术的结合为预测食用牛肝菌(Boletussect.Boletus)的地理来源建立了一套可靠的系统。